
- 4)Дыхательная цепь и комплексы с переносом заряда.
- 5)Структура и функции конкретных компонентов дыхательной цепи.
- Цепь переноса электронов (цпэ).
- Структурная организация цепи переноса электронов (цпэ).
- Компоненты цпэ:
- Строение дыхательной цепи (дц), комплексы, ингибиторы. Механизм работы. Пункты сопряжения, величина овп компонентов дц. Коэффициент р/о, его значение.
4)Дыхательная цепь и комплексы с переносом заряда.
Цепь переноса электронов митохондрий
Цепь переноса электронов (дыхательная цепь) – это сложная система переносчиков, при участии которой происходит процесс последовательного переноса электронов от NADH и FADH2 на O2.
Дыхательной цепи предшествует отнятие атомов водорода от окисляемых субстратов (реакции дегидрирования).
Эти реакции относятся к окислительно-восстановительным.
Их катализируют ферменты оксидоредуктазы (подкласс: дегидрогеназы).
В основе функционирования ЦПЭ лежит работа 5 основных компонентов, обеспечивающих перенос электронов от NADH и FADH2 на O2.
В их число входит 3 сложных ферментных комплексов, а также 2 низкомолекулярных переносчика.
NADH-дегидрогеназа (комплекс I);
сукцинатдегидрогеназа (комплекс II);
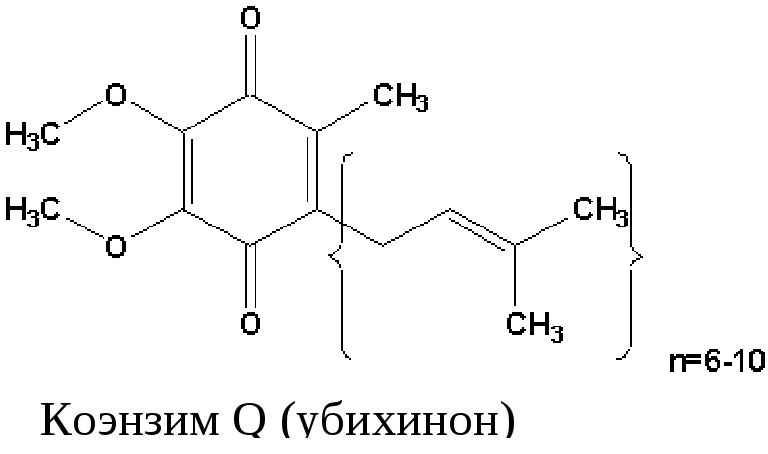
низкомолекулярный переносчик: убихинон (кофермент Q);
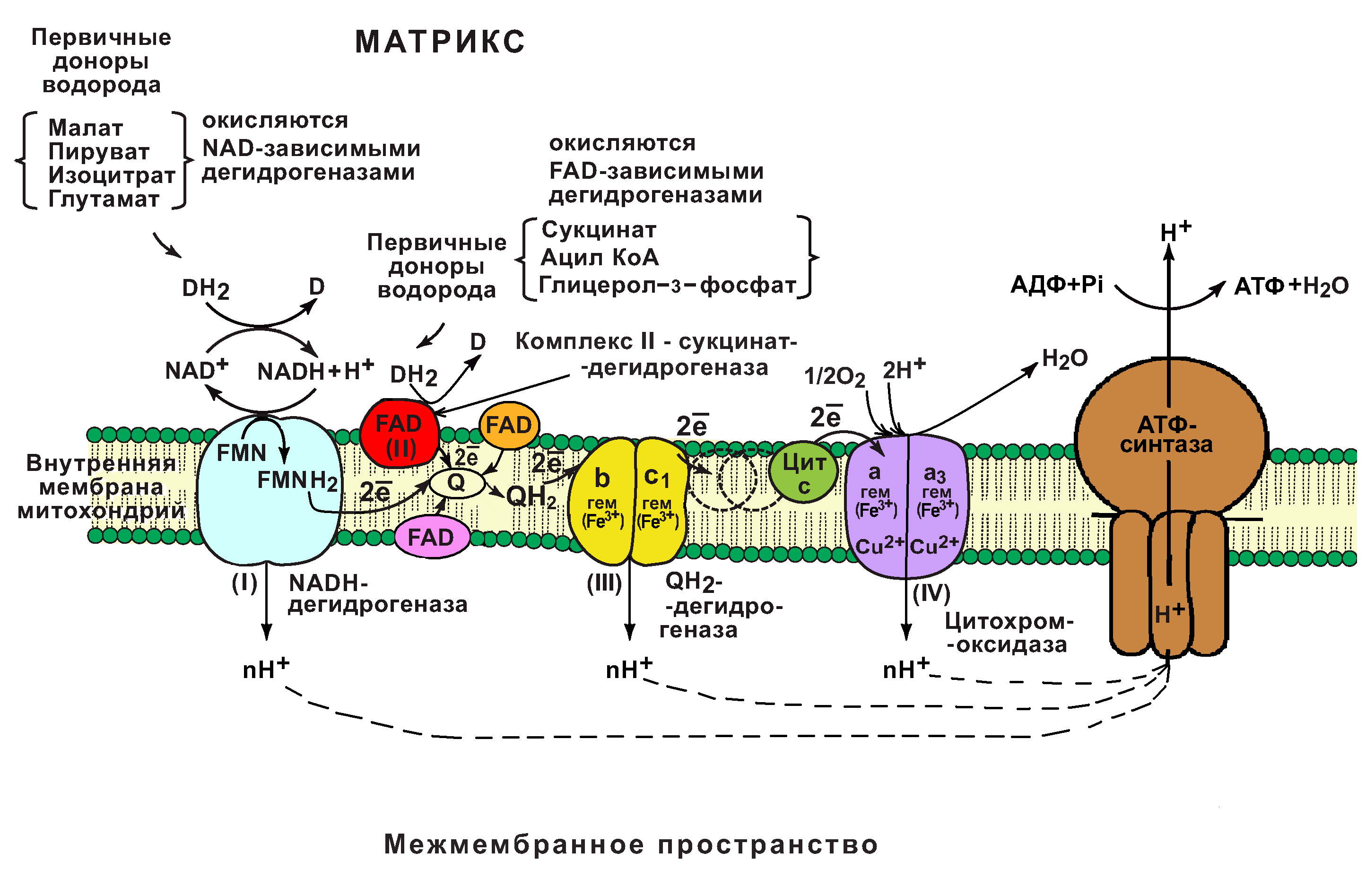 Убихинолдегидрогеназа (QН2-дегидрогеназа) (комплекс III);
Убихинолдегидрогеназа (QН2-дегидрогеназа) (комплекс III);
низкомолекулярный переносчик (цитохром с);
цитохромоксидаза (комплекс IV)
Все компоненты ЦПЭ располагаются во внутренней мембране митохондрий в порядке возрастания окислительно-восстановительных потенциалов (редокс-потенциалов).
Комплексы дыхательной цепи
Комплекс I (НАДН-дегидрогеназный комплекс) окисляет НАД-Н, отбирая у него два электрона и перенося их на растворимый в липидах убихинон, который внутри мембраны диффундирует к комплексу III. Вместе с этим, комплекс I перекачивает 2 протона и 2 электрона из матрикса в межмембранное пространство митохондрии.
Комплекс II (Сукцинатдегидрогеназа) не перекачивает протоны, но обеспечивает вход в цепь дополнительных электронов за счёт окисления сукцината.
Комплекс III (Цитохром-bc1-комплекс) переносит электроны с убихинона на два водорастворимых цитохрома с, расположенных на внутренней мембране митохондрии. Убихинон передаёт 2 электрона, а цитохромы за один цикл переносят по одному электрону. При этом туда также переходят 2 протона убихинона и перекачиваются комплексом.
Комплекс IV (Цитохром c оксидаза) катализирует перенос 4 электронов с 4 молекул цитохрома на O2 и перекачивает при этом 4 протона в межмембранное пространство. Комплекс состоит из цитохромов a и a3, которые, помимо гема, содержат ионы меди.
Кислород, поступающий в митохондрии из крови, связывается с атомом железа в гемецитохрома a3 в форме молекулы O2. Каждый из атомов кислорода присоединяет по два электрона и два протона и превращается в молекулу воды.
5)Структура и функции конкретных компонентов дыхательной цепи.
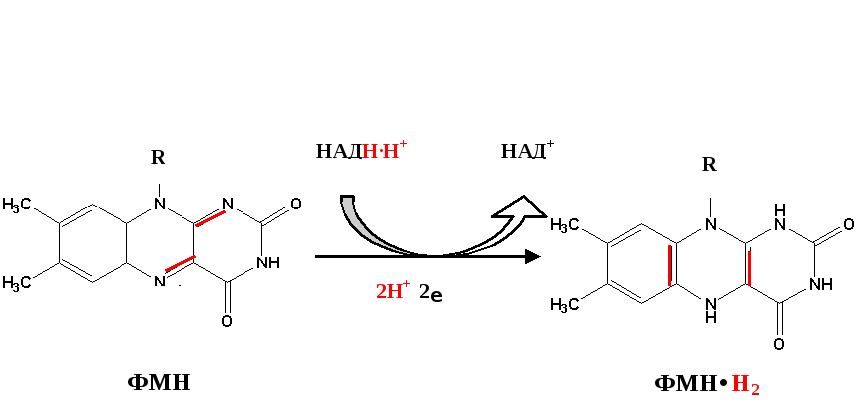
1) НАДН-дегидрогеназа(НАДН-КоQ-оксидоредуктаза) – встроена во внутреннюю мембрану митохондрий. Это флавин-зависимый фермент, в состав которого входит витамин В2. Сложный, имеет 2 простетические группы: ФМН (флавинмононуклеотид – активная форма витамина В2) и Железосерные белки (FеS-белки). АтомыFe, входящие в FеS-центры являются негеминовыми (не входят в состав гема). Активный центр НАДНДГ обращён в сторону матрикса митохондрий. НАДНДГ дегедрирует НАДН (отщепляет Н2) и передаёт протоны и электроны сначала на ФМН, а потом ч/з FеS-центры на СоQ(восстанавливается и превращается в КоQН2 или убихинол).
Функция: а)Принимает электроны от НАДН и передает их на коэнзим Q (убихинон).
б) Переносит 4 иона Н + на наружную поверхность внутренней митохондриальной мембраны.
2) КоQ (убихинон) – небелковый переносчик, находится в растворенном состоянии, т.к. является жирорастворимым (растворяется в липидном слое мембран). Поэтому может перемещаться как вдоль так и поперек мембраны (не заряжен). Поэтому он принимает электроны и протоны с ФАДН2, т.е. ч/з 2й вход в дыхательную цепь. Его называют коллектором электронов.
3) Цитохромы (b, c1, c, а, а3)– сложные белки (гемпротеины), небелковой частью которых является гем, содержащийFе 3+ (окисленная форма). Fегема может обратимо принимать и отдавать электроны.
4) УбихинолДГ (КоQ-цитохром с-оксидоредуктаза)– это ферментный комплекс, состоящий из цитохромовbи с1, включающий также FеS-центры. Функция:
а) Принимает электроны от коэнзима Q и передает их на цитохром с (при этом Fе восстанавливается до 2х валентного); б)Переносит 2 иона Н + на наружную поверхность внутренней митохондриальной мембраны.
5) Цитохром с-кислород-оксидоредуктаза — В этом комплексе находятся цитохромы а и а3, он называется также цитохромоксидаза, всего содержит 6 полипептидных цепей. В комплексе также имеется 2 иона меди. Функция: а) Принимает электроны от цитохрома с и передает их на кислород с образованием воды. б)Переносит 4 иона Н + на наружную поверхность внутренней митохондриальной мембраны.
6) АТФ-синтаза– комплекс, состоящий из множества белковых цепей, подразделенных на две большие группы: одна группа формирует субъединицу Fо (произносится со звуком «о», а не «ноль» т.колигомицин-чувствительная) – ее функция каналообразующая, по ней выкачанные наружу протоны водорода устремляются в матрикс. Другая группа образует субъединицу F1 – ее функция каталитическая, именно она, используя энергию протонов, синтезирует АТФ.
Источник
Цепь переноса электронов (цпэ).
Цепь переноса электронов (дыхательная цепь) – это сложная система переносчиков, при участии которой происходит процесс последовательного переноса электронов от NADH и FADH2 на O2.
Дыхательной цепи предшествует отнятие атомов водорода от окисляемых субстратов (реакции дегидрирования).
Эти реакции относятся к окислительно-восстановительным.
Их катализируют ферменты оксидоредуктазы (подкласс: дегидрогеназы).
Бóльшая часть окислительно-восстановительных реакций в клетке происходит в матриксе митохондрий.
Реакции дегидрирования происходят в ходе специфических и общего пути катаболизма.
Выделяют 2 типа дегидрогеназ, катализирующие реакции дегидрирования, предшествующие ЦПЭ:
NAD + – зависимые дегидрогеназы (предшественник: витамин PP);
! Субстратов NAD + – зависимых дегидрогеназ большинство.
Примеры: малат, изоцитрат, -кетоглутарат, глутаминовая кислота и др.
Кофермент NAD + не образует прочной постоянной связи с ферментом.
Он соединяется с АЦ фермента уже в ходе реакции и после её завершения уже в восстановленной форме отсоединяется.
FAD – зависимые дегидрогеназы (предшественник: витамин B2)
Кофермент FAD прочно связан с активным центром фермента как в окисленной, так и в восстановленной форме.
В ключаясь в реакцию, он принимает 2H + и 2 и
ключаясь в реакцию, он принимает 2H + и 2 и
переходит в восстановленную форму.
Субстраты FAD-зависимых дегидрогеназ: сукцинат, ацил-KoA, глицерол-3-фосфат (-глицерофосфат).
Структурная организация цепи переноса электронов (цпэ).
В основе функционирования ЦПЭ лежит работа 5 основных компонентов, обеспечивающих перенос электронов от NADH и FADH2 на O2.
В их число входит 3 сложных ферментных комплексов, а также 2 низкомолекулярных переносчика.
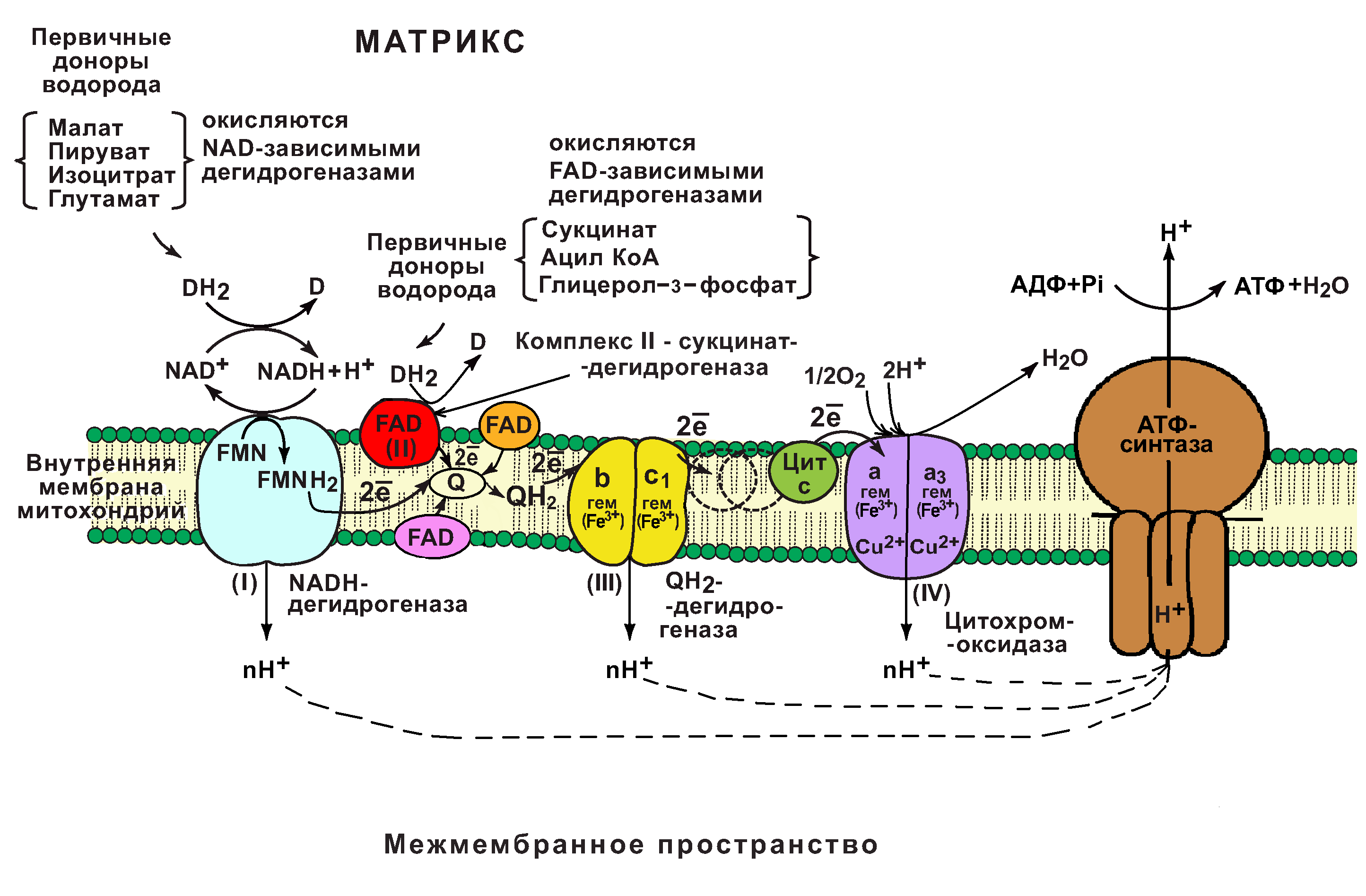
Компоненты цпэ:
NADH-дегидрогеназа (комплекс I);
сукцинатдегидрогеназа (комплекс II);
низкомолекулярный переносчик: убихинон (кофермент Q);
Убихинолдегидрогеназа (QН2-дегидрогеназа) (комплекс III);
низкомолекулярный переносчик (цитохром с);
цитохромоксидаза (комплекс IV)
Все компоненты ЦПЭ располагаются во внутренней мембране митохондрий в порядке возрастания окислительно-восстановительных потенциалов (редокс-потенциалов).
Самый низкий редокс-потенциал у NADH (-0,32В), а самый высокий редокс-потенциал у кислорода (+0,82В).
Это обеспечивает последовательное перемещение электронов от NADH (или FADH2) на O2, при котором происходит выделение энергии на каждом этапе ЦПЭ.
ЦПЭ обеспечивает взаимосвязь (сопряжение) процессов окисления и фосфорилирования АДФ.
Основная роль в переносе электронов принадлежит 3 важнейшим ферментным комплексам (I, III, IV).
Это сложные олигомерные белки, расположенные во внутренней мембране митохондрий.
Источник
Строение дыхательной цепи (дц), комплексы, ингибиторы. Механизм работы. Пункты сопряжения, величина овп компонентов дц. Коэффициент р/о, его значение.
Поэтапное «контролируемое сгорание» достигается путём промежуточного включения дыхательных ферментов, обладающих различным редокс-потенциалом. Редокс-потенциал (окислительно-восстановительный потенциал) определяет направление переноса протонов и электронов ферментами дыхательной цепи (рис.1).
Редокс-потенциал выражается значением электродвижущей силы (в вольтах), которая возникает в растворе между окислителем и восстановителем, присутствующих в концентрации 1,0 моль/л при 25˚ С (при рН=7,0 оба находятся в равновесии с электродом, который может обратимо принимать электроны от восстановителя). При рН=7,0 редокс-потенциал системы Н2 /2Н + +2ē равен – 0,42 v. Знак – означает, что данная редокс-пара легко отдаёт электроны, т.е. играет роль восстановителя, знак + указывает на способность редокс-пары принимать электроны, т.е. играть роль окислителя. Например, редокс-потенциал пары НАДН∙Н + / НАД + равен – 0,32 v, что говорит о высокой её способности отдавать электроны, а окислительно-восстановительная пара ½О2 /Н2О имеет наибольшую положительную величину +0,81 v, т.е. кислород обладает наивысшей способностью принимать электроны.
В процессе окисления АцКоА в ЦТК, восстановленные формы НАДН2 и ФАДН2 поступают в ДЦ, где энергия электронов и протонов трансформируется в энергию макроэргических связей АТФ.
ДЦ — совокупность дегидрогеназ, которые транспортируют электороны и протоны с субстрата на кислород.
Принципы функционирования ДЦ основаны на 1-ом и 2-ом законах термодинамики.
Движущей силой ДЦ является разность ОВП. Суммарная разность всей ДЦ составляет 1,1 В. Пункты фосфорилирования должны иметь перепад ОВП = 0,25 — 0,3 В.
1. Пара НАД-Н имеет ОВП = 0,32 В.
ДЦ локализуется во внутренней мембране митохондрий и имеет 2 пути введения электронов и протонов или 2 входа; ДЦ образует 4 комплекса.
1 вход: НАД-зависимый (поступают электроны и протоны со всех НАД-зависимых реакций).
Дыхательная цепь – форма реализации биологического окисления.
Тканевое дыхание – это последовательность окислительно-восстанови-тельных реакций, протекающих во внутренней митохондриальной мембране с участием ферментов дыхательной цепи. Дыхательная цепь имеет чёткую структурную организацию, её компоненты формируют дыхательные комплексы, порядок расположения которых зависит от величины их редокс-потенциала (рис.5.1). Количество дыхательных цепей в отдельно взятой митохондрии из клеток разных тканей неодинаково: в печени – 5000, в сердце – около 20 000, следовательно, миокардиоциты отличаются более интенсивным дыханием, чем гепатоциты.
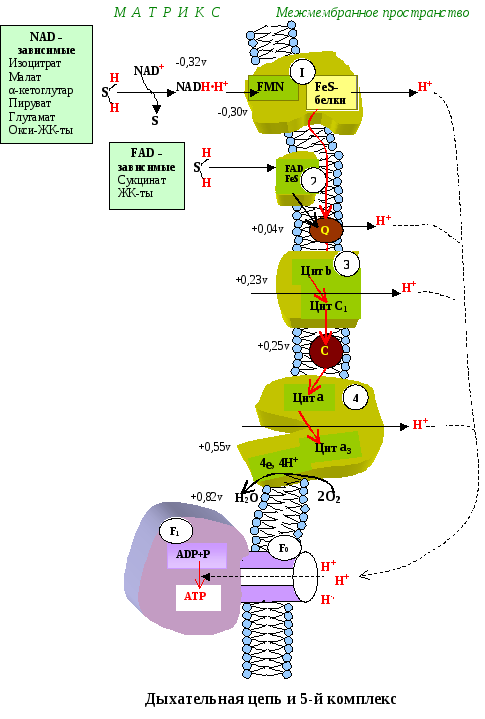
Рис. 5.1 Порядок расположения комплексов дыхательной цепи во внутренней мембране митохондрий
Прежде чем остановиться на характеристике каждого из компонентов дыхательной цепи, познакомимся с субстратами тканевого дыхания.
Субстраты тканевого дыхания подразделяются на 2 группы:
НАД-зависимые – субстраты цикла Кребса изоцитрат, α-кетоглутарат и малат. Это также пируват, гидроксибутират и β–гидрокси-ацил
КоА, глутамат и некоторые другие аминокислоты. Водород от НАД-зависимых субстратов c помощью НАД-зависимых дегидрогеназ передаётся на I-й комплекс дыхательной цепи.
ФАД-зависимые – сукцинат, глицерол-3-фосфат, ацил
КоА и некоторые другие. Водород от ФАД-зависимых субстратов передаётся на II-й комплекс дыхательной цепи.
При дегидрировании субстратов НАД-зависимыми дегидрогеназами образуется восстановленная форма НАД (НАДH∙H + ).
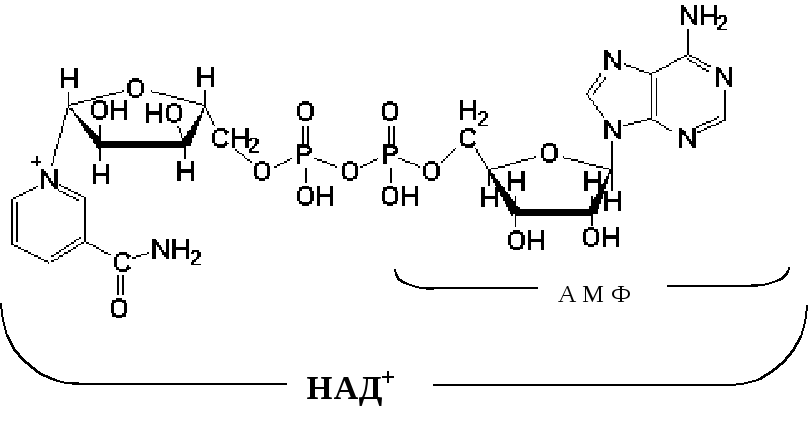
Указана окисленная форма кофермента НАД + . Этот кофермент является динуклеотидом (никотинамид—аденин—динуклеотид): в состав одного нуклеотида входит витамин РР (никотинамид), другой представляет собой АМФ. Способность кофермента играть роль промежуточного переносчика водородов связана с наличием в его структуре витамина РР. В электронно-протонной форме процесс обратимого гидрирования-дегидрирования может быть представлен уравнением (R- остальная часть кофермента):
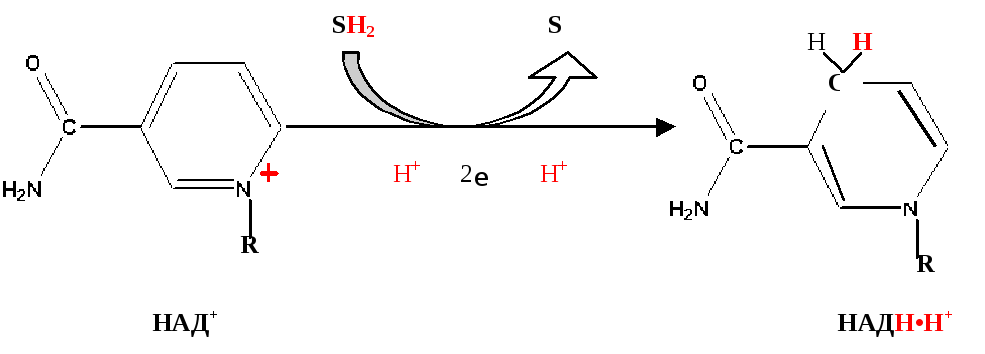
НАДH∙H + может образовываться не только в митохондриях, но и в цитозоле клетки при протекании определённых процессов метаболизма. Однако цитоплазматический кофермент не может проникать в митохондрии. Водород восстановленного кофермента должен быть сначала перенесен на субстраты, которые могут проникать в митохондрии. Такими «Н2-переносящими субстратами» являются:
Дигидроксиацетон фосфат → глицерол-3-фосфат
НАДH∙H + затем окисляется 1-м комплексом дыхательной цепи. Рассмотрим работу этого комплекса.
I комплекс цепи тканевого дыхания – НАДH∙H + -убихинон-оксидодуктаза.
Первый комплекс является самым большим в дыхательной цепи (представлен 23-30 субъединицами). Он катализирует перенос водорода от НАДH∙H + на убихинон (рис. 5.1 и рис. 5.3). В его состав входят кофермент ФМН (флавинмононуклеотид) и железосерные белки, содержащие негеминовое железо. Функция этих белков заключается в разделении потока протонов и электронов: электроны переносятся от ФМН∙Н2 к внутренней поверхности внутренней мембраны митохндрий (обращенной к матриксу), а протоны – к внешней поверхности внутренней мембраны и затем высвобождаются в митохондриальный матрикс.
При транспорте протонов и электронов редокс-потенциал первого комплекса снижается на 0,38 v, что вполне достаточно для синтеза АТФ. Однако в самом комплексе АТФ не образуется, а высвобождающаяся в результате работы комплекса энергия аккумулируется (см. ниже образование электро-химического потенциала) и частично рассеивается в виде тепла.
По своему строению ФМН – мононуклеотид, в котором азотистое основание представлено изоаллоксазиновым ядром рибофлавина, а пентозой является рибитол (иными словами, ФМН – это фосфорилированная форма витамина В2).
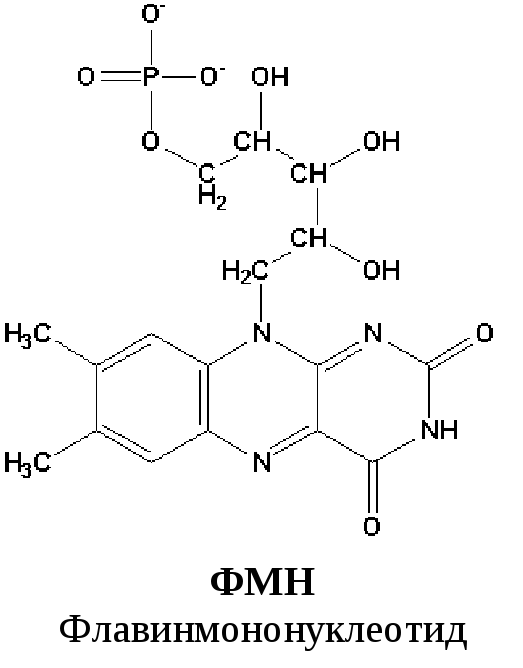
Функция ФМН заключается в акцепции 2 атомов водорода от НАДH∙H + и передачи их железосерным белкам. Водород (2 электрона и 2 протона) присоединяется к атомам азота изоаллоксазинового кольца, при этом происходит внутримолекулярная перегруппировка двойных связей с образованием промежуточного семихинона – соединения свободнорадикальной природы (на схеме представлено суммарное уравнение реакции, где R – остальная часть молекулы)
II комплекс цепи тканевого дыхания – сукцинат-убихинон-оксидоредуктаза.
Этот комплекс имеет меньшую молекулярную массу и также содержит железосерные белки. Сукцинат-убихинон-оксидоредуктаза катализирует перенос водорода от сукцината на убихинон. В состав комплекса входит кофермент ФАД (флавин-аденин-динуклеотид) и фермент сукцинатдегидрогеназа, который является одновременно ферментом цикла Кребса. Ацил
SКоА, 3-фосфо-глицерат и диоксиацетон фосфат также являются ФАД-зависимыми субстратами тканевого дыхания и с помощью этого кофермента контактируют со вторым комплексом.
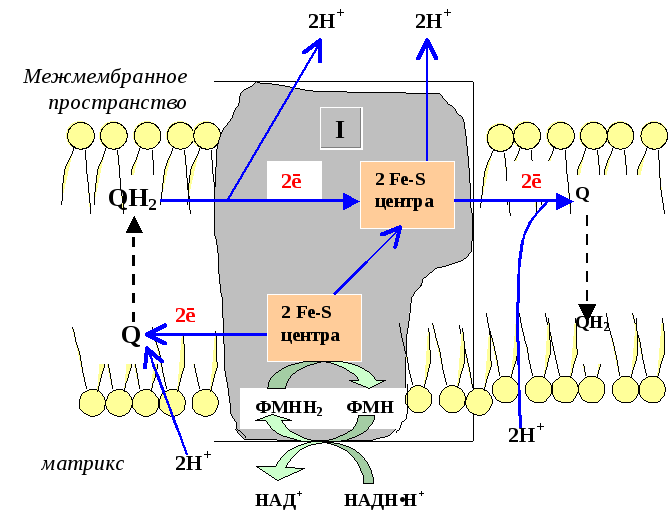
Рис. 5.3 Первый комплекс дыхательной цепи
Энергия включения водорода субстратов во II комплекс цепи тканевого дыхания рассеивается в основном в виде тепла, так как на этом участке цепи редокс-потенциал снижается незначительно и этой энергии для синтеза АТФ мало.
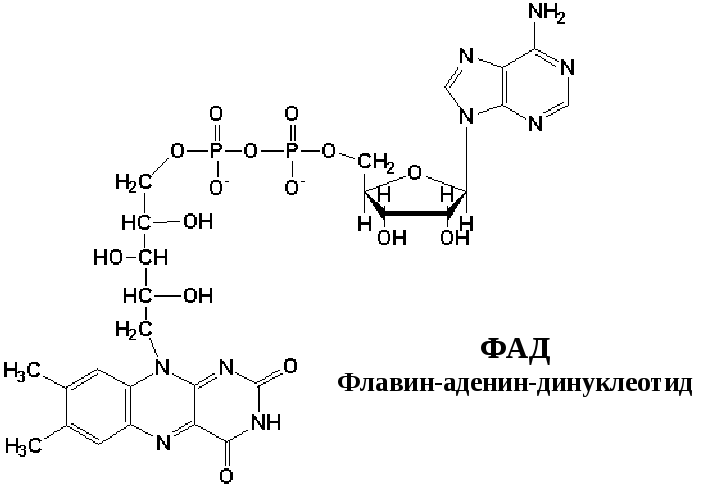
Процесс восстановления ФАД протекает аналогично таковому ФМН.
Кофермент Q или убихинон — гидрофобное соединение, является компонентом клеточных мембран, содержится в большой концентрации, относится к группе витаминов. относится к группе витаминов.
Убихинон (коэнзим Q). Убихинон – небольшая липофильная молекула, по химическому строению представляющая собой бензохинон с длинной боковой цепью (число изопреноидных единиц колеблется от 6 у бактерий до 10 у млекопитающих).
В дыхательной цепи коэнзим Q является своеобразным депо (пулом) водорода, который он получает от различных флавопротеинов. Липофильный характер молекулы убихинона обуславливает его способность свободно перемещаться в липидной фазе митохондриальной мембраны, перехватывая протоны и электроны не только от I и II комплексов дыхательной цепи, но и захватывая из митохондриального матрикса протоны. При этом убихинон восстанавливается с образованием промежуточного свободнорадикального продукта – семихинона .
Восстановленная форма убихинона – убихинол – передаёт протоны и электроны на III комплекс дыхательной цепи.
Цитохромоксидаза имеет высокую степень сродства к кислороду и может работать при его низких концентрациях.
аа3 — состоит из 6 субъединиц каждая из которых содержит гем и атом меди. 2 субъединицы составляют цитохром а, а остальные 4 относятся к цитохрому а3.
Между НАД и ФП, b-c, a-a3 имеет место max перепад ОВП. Эти пункты являются местом синтеза АТФ (местом фосфорилирования АДФ).
III комплекс цепи тканевого дыхания – убихинол-цитохром С-оксидоредуктаза. В состав III комплекса входят цитохромы b и с1, относящиеся к группе сложных белков хромопротеинов. Простетическая группа этих белков окрашена (chroma – краска) и близка по химическому строению к гему гемоглобина. Однако в противоположность гемоглобину и оксигемоглобину, в которых железо должно быть только в 2-х валентной форме, железо в цитохромах при работе дыхательной цепи переходит от двух- к трёхвалентному состоянию (и обратно).
Как видно из названия, III комплекс переносит электроны от убихинола на цитохром С. Вначале электроны поступают на окисленную форму цитохрома b (Fe 3+ ), который при этом восстанавливается (Fe 2+ ), затем восстановленный цитохром b передаёт электроны окисленной форме цитохрома с, который также восстанавливается и, в свою очередь, передаёт электроны цитохрому С.
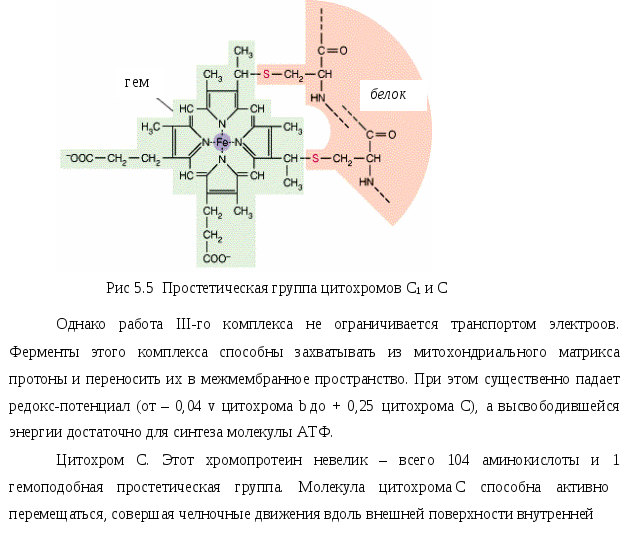
митохондриальной мембраны от III комплекса к IV и обратно. При этом 1 молекула цитохрома С, попеременно окисляясь и восстанавливаясь, переносит 1 электрон.
IV комплекс дыхательной цепи – цитохром С-оксидаза. Комплекс назван оксидазой из-за способности непосредственно взаимодействовать с кислородом. У млекопитающих этот крупный (
200 kD) трансмембранный белок состоит из 6-13 субъединиц, из которых некоторые кодируются митохондриальной ДНК. В состав IV комплекса входят 2 хромопротена – цитохром а и цитохром а3. В отличие от других цитохромов, цитохромы а и а3 каждый содержат не только атом железа, но и атом меди. Медь в составе этих цитохромов при транспорте электронов также попеременно переходит в окисленное (Cu 2+ ) и восстановленное (Cu + ) состояние.
Цитохром с-оксидаза катализирует одноэлектронное окисление 4-х восстановленных молекул цитохрома с и при этом одновременно осуществляет полное (4-х электронное) восстановление молекулы кислорода:
4  цитохрома с (Fe 2+ ) + 4 H + + O2 4 цитохрома с (Fe 3+ ) + H2O
цитохрома с (Fe 2+ ) + 4 H + + O2 4 цитохрома с (Fe 3+ ) + H2O
Протоны для образования молекул воды поступают из матрикса. Следует заметить, что эта реакция весьма сложна и протекает через промежуточные стадии образования свободных радикалов кислорода.
Окислительно-восстановительный потенциал IV комплекса является самым большим (+0,57 v), его энергии вполне достаточно для синтеза 3-х молекул АТФ, однако большая часть этой энергии используется на «перекачивание» протонов из матрикса митохондрий в межмембранное пространство. В связи с активным транспортом протонов цитохром с-оксидаза получила название «протонного насоса».
Таким образом, тканевое дыхание представляет собой процесс транспорта электронов и протонов от НАД- или ФАД-зависимых субстратов на кислород, а также протонов, поставляемых матриксом митохондрий. При транспорте падает редокс-потенциал, что сопровождается высвобождением заключённой в субстратах тканевого дыхания энергии. Полное восстановление молекулярного кислорода воздуха в дыхательной цепи сопровождается образованием воды.
Источник