
- БИОХИМИЯ — Л. Страйер — 1984
- ТОМ 3
- Часть IV ИНФОРМАЦИЯ
- ГЛАВА 24 ДНК: ГЕНЕТИЧЕСКАЯ РОЛЬ, СТРУКТУРА И РЕПЛИКАЦИЯ
- 24.19. Одна цепь ДНК синтезируется прерывисто
- 24.20. Затравкой синтеза ДНК служит РНК
- 24.21. Энергия гидролиза АТР используется для расплетания родительской ДНК в области репликационной вилки под действием белка rep
- 24.22. ДНК-гираза вводит отрицательные супервитки в родительскую ДНК, чтобы облегчить ее расплетание
- 9. Нуклеиновые кислоты. Строение и функции днк. Генетический код, его структура и свойства.
- 10. Воспроизведение на молекулярном уровне. Репликация днк. Понятие и репарации днк.
БИОХИМИЯ — Л. Страйер — 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 24 ДНК: ГЕНЕТИЧЕСКАЯ РОЛЬ, СТРУКТУРА И РЕПЛИКАЦИЯ
24.19. Одна цепь ДНК синтезируется прерывисто
Вернемся к взаимодействию молекул при репликации. В области репликационной вилки обе цепи родительской ДНК служат матрицами для синтеза новой ДНК. Напомним, что цепи родительской ДНК антипараллельны. Следовательно, общее направление синтеза ДНК должно быть 5′ → 3′ для одной из дочерних цепей и 3′ → 5′ для другой (рис. 24.39). Однако все известные ДНК-полимеразы синтезируют ДНК в направлении 5′ → 3′, а не 3′ → 5′. Как же тогда происходит кажущийся (при низкой разрешающей способности метода) рост одной из дочерних цепей в направлении 3′ → 5′?
Рис. 24.39. При низком разрешении кажущееся направление репликации ДНК будет 5′ → 3′ для одной дочерней цепи и 3′ → 5′ для другой. На самом деле обе цепи синтезируются в направлении 5′ → 3′, как показано на рис. 24.40

Проблема была решена Рейдзи Оказаки (Reiji Okazaki), который обнаружил, что значительная часть новосинтезированной ДНК существует в виде коротких фрагментов. Такие фрагменты длиной около 1000 нуклеотидов (они называются фрагментами Оказаки) существуют в течение непродолжительного времени в непосредственной близости от репликационной вилки. По мере прохождения репликации эти фрагменты соединяются друг с другом ковалентно под действием ДНК-лигазы и образуют одну из дочерних цепей (рис. 24.40). Другая новая цепь синтезируется непрерывно или почти непрерывно. Та цепь, которая образуется из фрагментов Оказаки, называется отстающей цепью, а та, что синтезируется без разрывов или почти без разрывов, ведущей цепью. И фрагменты Оказаки, и ведущая цепь синтезируются в направлении 5′ → 3′. Прерывистая сборка отстающей цепи позволяет путем полимеризации в направлении 5′ → 3′ на атомном уровне получать общий рост цепи в направлении 3′ → 5′.
Рис. 24.40. Схематическое изображение репликационной вилки. Обе цепи ДНК синтезируются в направлении 5′ → 3′. Ведущая цепь синтезируется непрерывно, а отстающая — в виде коротких фрагментов (фрагменты Оказаки)

24.20. Затравкой синтеза ДНК служит РНК
Как начинается синтез ДНК? Напомним, что всем ДНК-полимеразам для инициирования синтеза ДНК необходима затравка со свободной З’-ОН-группой. Что служит затравкой при синтезе ведущей цепи и фрагментов Оказаки? Важным толчком к решению этого вопроса послужило наблюдение, что для инициации синтеза ДНК необходим синтез РНК. На основе этого открытия было высказано предположение, что РНК, очевидно, служит затравкой в синтезе ДНК, так как уже было известно, что РНК-полимеразы способны начинать синтез цепей de novo. Затем было показано, что новообразующаяся ДНК ковалентно связана с коротким фрагментом РНК, который и служит затравкой. Итак, РНК-затравка в синтезе ДНК.
По всей вероятности, репликация ДНК в клетке E. coli происходит, как показано на рис. 24.41.
Рис. 24.41. Инициация синтеза ДНК. А — праймаза синтезирует короткую комплементарную цепь РНК; Б — эта РНК служит затравкой для синтеза новой ДНК; В — РНК, входящая в состав новообразованной цепи, гидролизуется, при этом образуется брешь, которая впоследствии заполняется

1. Особая РНК-полимераза (ее называют праймаза) синтезирует короткую цепь РНК (примерно 10 нуклеотидов), комплементарную одной из цепей ДНК-матрицы. В отличие от ДНК-полимеразы праймаза не нуждается в затравке для синтеза полинуклеотида.
2. 3′-гидроксильная группа концевого рибонуклеотида этой цепи РНК служит затравкой для синтеза ДНК под действием голофермента ДНК-полимеразы III. Большая часть новообразованной ДНК синтезируется этим мультисубъединичным комплексом.
3. РНК-компонент этого РНК-ДНК-гибрида гидролизуется под действием ДНК- полимеразы I.
4. После удаления РНК из новообразованных цепей между фрагментами ДНК остаются довольно обширные бреши. ДНК- полимераза I, которая хорошо приспособле-
на для синтеза ДНК на одноцепочечной матрице, заполняет эти бреши.
Последние исследования показали, что действию праймазы предшествует образование предзатравочного промежуточного комплекса, состоящего как минимум из пяти белков. Один из них — белок dnaB — может передвигаться вдоль ДНК, используя энергию гидролиза АТР. Белок dnaB может служить сигналом для активации праймазы. Специфичность инициации очередного цикла репликации может обеспечиваться белками, доставляющими белок dnaB точно в область гена ilv, где находится точка начала репликации хромосомы E. coli. Время начала репликации ДНК имеет критическое значение, поскольку оно должно быть скоординировано с делением клетки. И действительно, бактериальная хромосома ассоциирована с впячиванием клеточной мембраны (рис. 24.42).
Рис. 24.42. Электронная микрофотография хромосомы Е. coli, прикрепленной к двум фрагментам клеточной мембраны. На рисунке изображена одна интактная суперспирализованная молекула ДНК

24.21. Энергия гидролиза АТР используется для расплетания родительской ДНК в области репликационной вилки под действием белка rep
В 1953 г. Уотсон и Крик отмечали, что «раскрутить спираль — задача труднопреодолимая». Исследования, проведенные в последнее время, показали, что в клетке E. coli в области репликационной вилки происходит активное расплетание родительской двойной спирали под действием фермента — белка rep. Поскольку энергия для расплетания родительской ДНК высвобождается при гидролизе АТР, белок гер называют хеликазой. На разделение каждой пары оснований затрачиваются примерно две молекулы АТР. Затем каждая из разделенных цепей родительской ДНК взаимодействует с несколькими молекулами белка, связывающегося с одноцепочечной ДНК (ОЦ-связывающий белок). Роль ОЦ-связывающего белка — стабилизировать одноцепочечные участки ДНК, образовавшиеся под действием хеликазы, чтобы расплетенная область могла функционировать в качестве матрицы. ОЦ-связывающие белки называют также белками, дестабилизирующими спираль (белки ДС), или плавящими белками.
24.22. ДНК-гираза вводит отрицательные супервитки в родительскую ДНК, чтобы облегчить ее расплетание
При расплетании ковалентно замкнутой кольцевой молекулы ДНК возникают топологические проблемы, так как расплетание двойной спирали вызывает образование положительных супервитков в замкнутой молекуле. В области репликационной вилки родительская ДНК вращается со скоростью 100 об/с — более чем в 100 раз быстрее, чем обычная долгоиграющая пластинка. Чтобы процесс расплетания продолжался, необходимо снять эти положительные супервитки, образовавшиеся в результате вращения. Другими словами, необходимо наличие какого-то молекулярного шарнира. Недавно Мартин Геллерт (Martin Gellert) установил, что эту функцию выполняет ДНК-гираза. Эта топоизомераза удаляет положительные супервитки, внося одноцепочечные разрывы и затем заделывая фосфодиэфирные связи в остове ДНК. АТР не требуется для такой термодинамически выгодной релаксации третичной структуры ДНК. Более того, ДНК-гираза может активно вводить отрицательные супервитки в ковалентно замкнутую кольцевую ДНК за счет энергии гидролиза АТР (рис. 24.43). Эти отрицательные супервитки способствуют расхождению цепей родительской ДНК в области репликационной вилки 1 .
1 В этой главе автор называет ДНК-гиразой два совершенно различных фермента. Один из них — ДНК-гираза, способная вводить в ДНК термодинамически невыгодные отрицательные супервитки за счет энергии гидролиза АТР; другой — независимая от АТР ДНК-топоизомераза, приводящая кольцевую молекулу ДНК в термодинамически равновесное (релаксированное) состояние. Это два совершенно различных белка: у них различные ингибиторы, потребности в ионных условиях среды, механизм действия. В репликации в качестве молекулярного шарнира участвует ДНК-гираза. Прим. перев.
Рис. 24.43. Каталитические активности ДНК-гиразы

Биологическая библиотека — материалы для студентов, учителей, учеников и их родителей.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
© 2018-2023 Все права на дизайн сайта принадлежат С.Є.А.
Источник
9. Нуклеиновые кислоты. Строение и функции днк. Генетический код, его структура и свойства.
ДНК состоит из нуклеотидов, в состав которых входят сахар – дезоксирибоза, фосфат и одно из азотистых оснований – пурин (аденин или гуанин) либо пиримидин (тимин или цитозин).
Молекулы ДНК включают в себя 2 полинуклеотидные цепи, соединенные друг с другом азотистыми основаниями с помощью водородных связей по принципу комплементарности (аденин -2вод.связи-тимин, гуанин-3вод.связи-цитозин).
Цепи антипараллельны: 5’-конец одной цепи соединяется с 3’-концом другой цепи. Чаще всего спирали правозакрученные.
В структурной организации молекулы ДНК можно выделить:
первичную структуру – полинуклеотидную цепь,
вторичную структуру – две комплементарные друг другу и антипараллельные полинуклеотидные цепи, соединенные водородными связями,
третичную структуру – трехмерную спираль с приведенными выше пространсвенными характеристиками.
Молекулы ДНК хранят (содержат) наследственную информацию (программу) о структуре специфических для каждого организма белков.
М 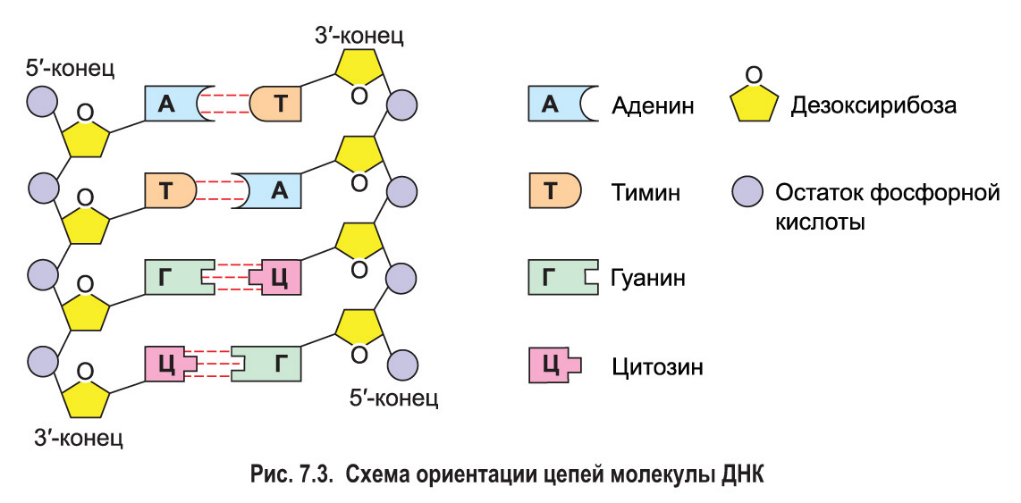 олекулы ДНК обеспечивают передачу наследственной информации от клетки к клетке, от организма к организму.
олекулы ДНК обеспечивают передачу наследственной информации от клетки к клетке, от организма к организму.
Молекулы ДНК участвуют в реализации генетической информации, т. е. участвуют в процессе синтеза полипептидов.
Генетический код – это система кодирования информации о последовательности аминокислот в белке с помощью последовательности нуклеотидов ДНК (РНК). Единицей генетического кода является кодон (триплет) – последовательность из трех нуклеотидов. В составе генетического кода 64 кодона, из них кодирующих аминокислоты – 61, а некодирующих (стоп-кодоны) – 3. Кодон АУГ, кодирующий аминокислоту метионин, выполняет функцию кодона-инициатора (старт-кодона) – с него начинается считывание генетической информации с иРНК. Первой к месту синтеза белка – рибосоме подходит тРНК с аминокислотой метионин. У эукариот только один старт-кодон, тогда как у прокариот их может быть несколько.
Свойства генетического кода:
триплетность – каждая аминокислота кодируется сочетанием из трех нуклеотидов;
специфичность – каждый кодон кодирует одну определенную аминокислоту;
вырожденность (избыточность) – почти каждая аминокислота (кроме метионина и триптофана) может быть закодирована двумя или более разными кодонами;
универсальность – принцип кодирования аминокислот у всех видов одинаков
10. Воспроизведение на молекулярном уровне. Репликация днк. Понятие и репарации днк.
Одна из основных функций ДНК – сохранение и передача наследственной информации. В основе этой функции лежит способность ДНК к самоудвоению – репликации. В результате репликации из одной материнской молекулы ДНК образуются две 10дочерние молекулы ДНК. Каждая дочерняя молекула содержит одну материнскую и одну вновь синтезированную цепь (полуконсервативный способ удвоения). Репликация в клетке происходит перед каждым её делением в синтетический период интерфазы (S-период).
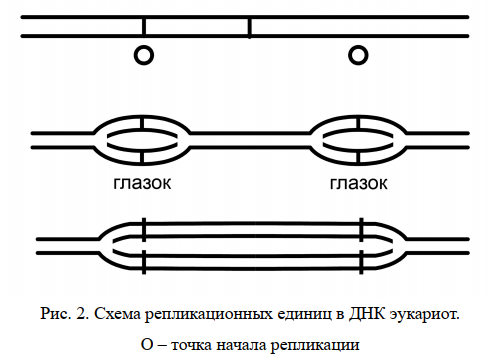
Единицей репликации является репликон – участок ДНК, способный к самостоятельной репликации. У бактерий и вирусов имеется обычно один репликон на клетку, тогда как у эукариот их содержится много. Репликация начинается в сайте инициации. Двойная спираль ДНК расплетается в точках инициации репликации и образуются репликационные глазки, состоящие из двух репликационных вилок, ведущих синтез в противоположных направлениях (двунаправленная репликация)
В синтезе полинуклеотидных цепей ДНК участвует фермент ДНК-полимераза, которая наращивает новую цепь в направлении 5′ → 3′. Однако синтез дочерних цепей ДНК осуществляется неодинаково. Одна из них создается непрерывно и называется лидирующей. Другая называется отстающей, так как она собирается из отдельных коротких участков (фрагментов Оказаки), которые потом соединяются вместе ДНК-лигазами. Такой феномен наблюдается из-за антипараллельности материнских цепей ДНК.
ФЕРМЕНТЫ и другие БЕЛКИ, обеспечивающие репликацию ДНК:
Геликаза – расплетает двойную спираль ДНК, разрушая водородные связи между комплементарными азотистыми основаниями.
РНК-полимераза (или ДНК-праймаза) – инициирует синтез ДНК, образуя РНК-затравки (праймеры).
ДНК-полимераза – синтезирует полинуклеотидную цепь ДНК в направлении 5′ → 3′.
ДНК-лигаза – сшивает вместе фрагменты Оказаки после удаления РНК-праймера и его замещения на нуклеотиды ДНК.
ДНК-топоизомераза – помогает раскручиванию ДНК и работе геликазы, снимая напряжение на спирали материнской молекулы.
Дестабилизирующие белки – негистоновые ядерные белки, связываются с разъединенными цепями ДНК, поддерживают репликационную вилку открытой.
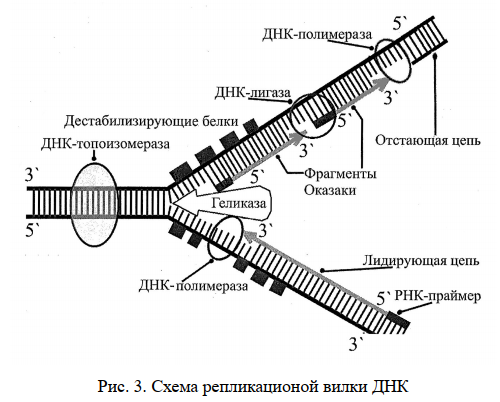
Геликаза — первый фермент, прикрепляется к точке начала репликации. Задача геликазы — продвигать вилки репликации вперед, «расплетая» ДНК (разрывая водородные связи между азотистыми основаниями).
Специальные белки, связывающиеся с одноцепочечной ДНК, обволакивают отдельные нити ДНК возле репликационной вилки, предотвращая их повторное объединение в двойную спираль.
ДНК-полимеразы могут добавлять нуклеотиды только к 3′-концу существующей цепи ДНК. Они используют свободную группу -ОН, находящуюся на 3′-конце, в качестве «крючка», прицепляя новый нуклеотид к этой группе в результате реакции полимеризации. Как же тогда ДНК-полимераза присоединяет первый нуклеотид на новой репликационной вилке?
В одиночку она это сделать не может! Эта проблема решается с помощью фермента праймазы. Праймаза создает РНК праймер или короткий фрагмент нуклеиновой кислоты, комплементарный матрице, который формирует 3′-конец для работы ДНК-полимеразы. Типичный праймер имеет длину от пяти до десяти нуклеотидов. Праймер способствует началу синтеза ДНК, то есть даёт ему старт.
Как только РНК-праймер синтезирован, ДНК-полимераза «удлинняет» его, добавляя нуклеотиды один за другим, чтобы получить новую цепь ДНК, которая комплементарна матричной цепи.
Лидирующая и отстающая цепи
У бактерии E. coli, ДНК-полимераза, которая осуществляет большую часть синтеза, является ДНК-полимеразой III. На репликационной вилке находятся две молекулы ДНК-полимеразы III, каждая из которых усердно работает над одной из двух новых цепей ДНК.
ДНК-полимеразы могут добавлять нуклеотиды только к 3′-концу существующей цепи ДНК. Они используют свободную группу -ОН, находящуюся на 3′-конце, в качестве «крючка», прицепляя новый нуклеотид к этой группе в результате реакции полимеризации. Как же тогда ДНК-полимераза присоединяет первый нуклеотид на новой репликационной вилке?
Одна новая цепь, которая проходит от 5′ к 3′-концу по направлению к вилке репликации, является простой. Эта цепь создается непрерывно, потому что ДНК-полимераза движется в том же направлении, что и вилка репликации. Эта непрерывно синтезируемая цепь называется лидирующей цепью.
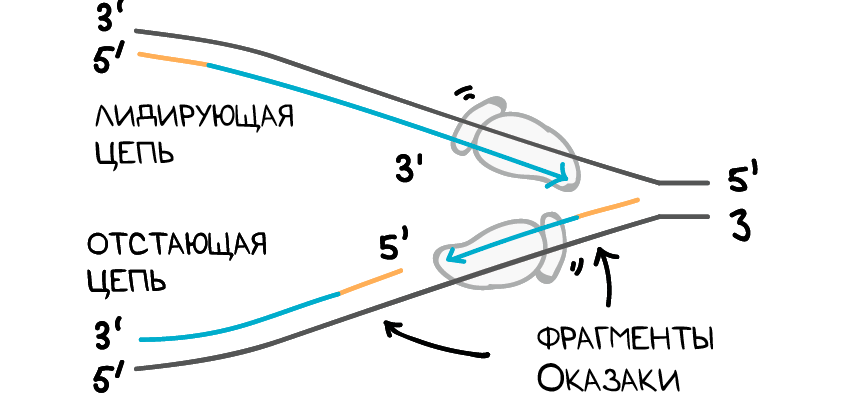
В другой новой цепи, которая направлена от репликационной вилки от 5′-конца к 3′-концу, всё немного сложнее. Эта цепь строится фрагментами, потому что по мере движения вилки ДНК-полимераза удаляется от неё, и ей в какой-то момент приходится отрываться и снова присоединяться к новому открывшемуся фрагменту. Эта цепь, достраивающаяся фрагментами, называется отстающей цепью.
Маленькие фрагменты называются фрагментами Оказаки. Они названы в честь японского ученого, который их обнаружил. Лидирующая цепь может быть синтезирована на основании одного праймера, в то время как отстающая цепь нуждается в новом праймере для каждого из коротких фрагментов Оказаки.
Бригада по техническому обслуживанию и уборке
Существуют и другие белки и ферменты, помимо перечисленных выше основных, необходимые для обеспечения бесперебойной репликации ДНК. В частности, это так называемые белки скользящего зажима, которые во время синтеза ДНК удерживают молекулы ДНК-полимеразы III. «Скользящий зажим» — это белок в форме кольца, не дающий ДНК-полимеразе III отсоединиться при переходе к новому фрагменту Оказаки^44start superscript, 4, end superscript.
Топоизомераза также играет важную поддерживающую роль в репликации ДНК. Этот фермент предотвращает слишком плотное скручивание двойной спирали ДНК перед вилкой репликации при раскрытии ДНК. Он вносит временные разрывы в спирали, чтобы снять излишнюю спирализацию цепей, а затем восстанавливает их, чтобы избежать необратимых повреждений.
Наконец, после всех этих процессов нужно сделать небольшую уборку, если мы хотим, чтобы в ДНК не оставалось РНК или зазоров. ДНК-полимераза I, ещё одна участвующая в репликации полимераза, удаляет РНК-праймеры и заменяет их на ДНК. Разрывы, которые остаются после удаления праймеров, восстанавливаются ферментом ДНК-лигазой.
Итоги: репликации ДНК у бактерии E. coli
Давайте уменьшим масштаб и посмотрим, как ферменты и белки, участвующие в репликации, работают вместе, чтобы синтезировать новую ДНК.
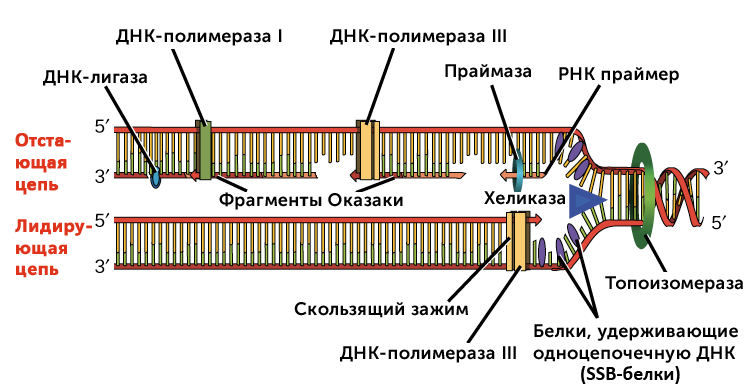
На рисунке показана репликационная вилка. Хеликаза разматывает спираль, а белки связывающие одноцепочечную ДНК предотвращают повторное формирование спирали. Топоизомераза предотвращает слишком плотную спирализацию ДНК перед репликационной вилкой. ДНК-праймаза создает РНК-праймер, а ДНК-полимераза достраивает цепь ДНК на основе РНК-праймера. Синтез ДНК происходит только в направлении от 5′-конца к 3′-концу. На лидирующей цепи синтез ДНК происходит непрерывно. На отстающей цепи синтез ДНК возобновляется много раз, по мере раскручивания спирали, что приводит к появлению множества коротких фрагментов, называемых «фрагментами Оказаки». ДНК-лигаза соединяет фрагменты Оказаки в единую молекулу ДНК.
Хеликаза раскрывает ДНК в репликационной вилке.
Белки, связывающие одноцепочечную ДНК (их также называют SSB-белки), удерживают разделённые цепи ДНК вблизи репликационной вилки, предотвращая их обратное соединение.
Топоизомераза работает перед вилкой репликации и предотвращает чрезмерное скручивание.
Праймаза синтезирует РНК праймеры комплементарные цепи ДНК.
ДНК полимераза III удлинняет праймеры, добавляя нуклеотиды к 3′-концу и создавая основную часть новой ДНК.
РНК праймеры удаляются и заменяются на ДНК с помощью ДНК полимеразы I.
Разрывы между фрагментами ДНК восстанавливаются ДНК лигазой.
Во время репликации, ДНК и рекомбинации ядерного материала в ней (сестринские хроматидные обмены, кроссинговер и др.) возможны «ошибки» в ДНК, и, как следствие этого, могут возникнуть мутации. Однако большинство «ошибок» и повреждений ДНК устраняется благодаря репарации – процессу, обеспечивающему исправление повреждений в ДНК.
Источник